RNA 干扰
概念
RNA 干扰(RNA interference, RNAi)是指在进化过程中高度保守的、由双链RNA(double-strand RNA, dsRNA)诱发的、同源mRNA高效特异性降解的现象,它是生物进化过程中遗留下来的一种在转录后通过RNA调控基因表达的机制。
原理
基因沉默分为转录水平的沉默(TGS)和转录后水平的沉默(PTGS)。TGS 是指转基因在细胞核内 RNA 合成受到了阻止而导致基因沉默;对于部分植物来说,转基因引发的基因沉默可能是因为基因特异的甲基化而导致;PTGS则是指转基因能够在细胞核里被稳定地转录,但在细胞质里却无相应的 mRNA 存在的这一现象。目前普遍认为 RNAi、共抑制、quelling 均属于 PTGS。
病毒基因、人工转入基因、转座子等外源性基因随机整合到宿主细胞基因内,并利用宿主细胞进行转录时,常产生一些 dsRNA。宿主细胞对这些 dsRNA 迅即产生反应,其胞质中的核酸内切酶 Dicer 将 dsRNA 切割成多个具有特定长度和结构的小片段 RNA(大约2123 bp),即 siRNA。研究表明在生物中 siRNA 具相似的结构特征:为长约2123 bp 的双链 RNA,具 5’单磷酸和 3’羟基末端,互补双链的 3’端均有一个2~3 nt 的单链突出。
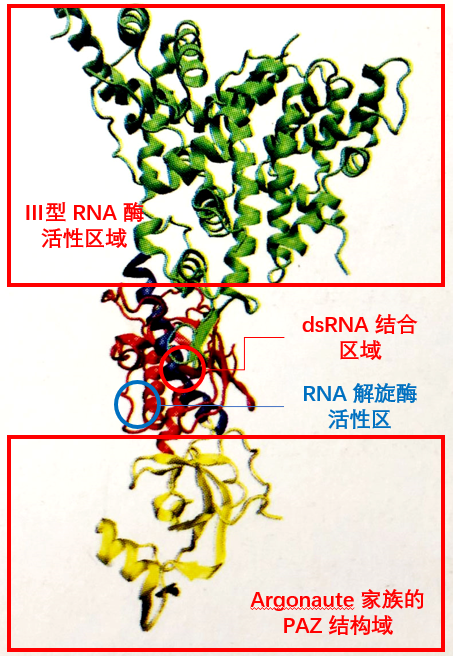
负责将 dsRNA 转化为 siRNA 的 Dicer核酸酶,属于 RNaseⅢ 家族,具有两个催化结构域、一个解旋酶(helicase)结构域和一个 PAZ (Piwi/Argonaute/Zwille)结构域,Dicer 在催化过程中以二聚体的形式出现,其催化结构域在 dsRNA 上反平行排列,形成四个活性位点,但只有两侧的两个位点有内切核酸酶活性,这两个位点在相距约 22 bp 的距离切断 dsRNA,各种生物体内 Dicer 结构略有不同,致使 siRNA 长度存在微小差别。
siRNA 在细胞内 RNA 解旋酶的作用下解链成正义链和反义链,继之由反义 siRNA 再与体内一些酶(包括内切酶、外切酶、解旋酶等)结合形成 RNA诱导的沉默复合物(RNA-induced silencing complex, RISC)。RISC 与外源性基因表达的mRNA的同源区进行特异性结合,RISC 具有核酸酶的功能,在结合部位切割 mRNA,导致特定基因沉默,切割位点即是与 siRNA 中反义链互补结合的两端。被切割后的断裂 mRNA 随即降解,从而诱发宿主细胞针对这些 mRNA 的降解反应。
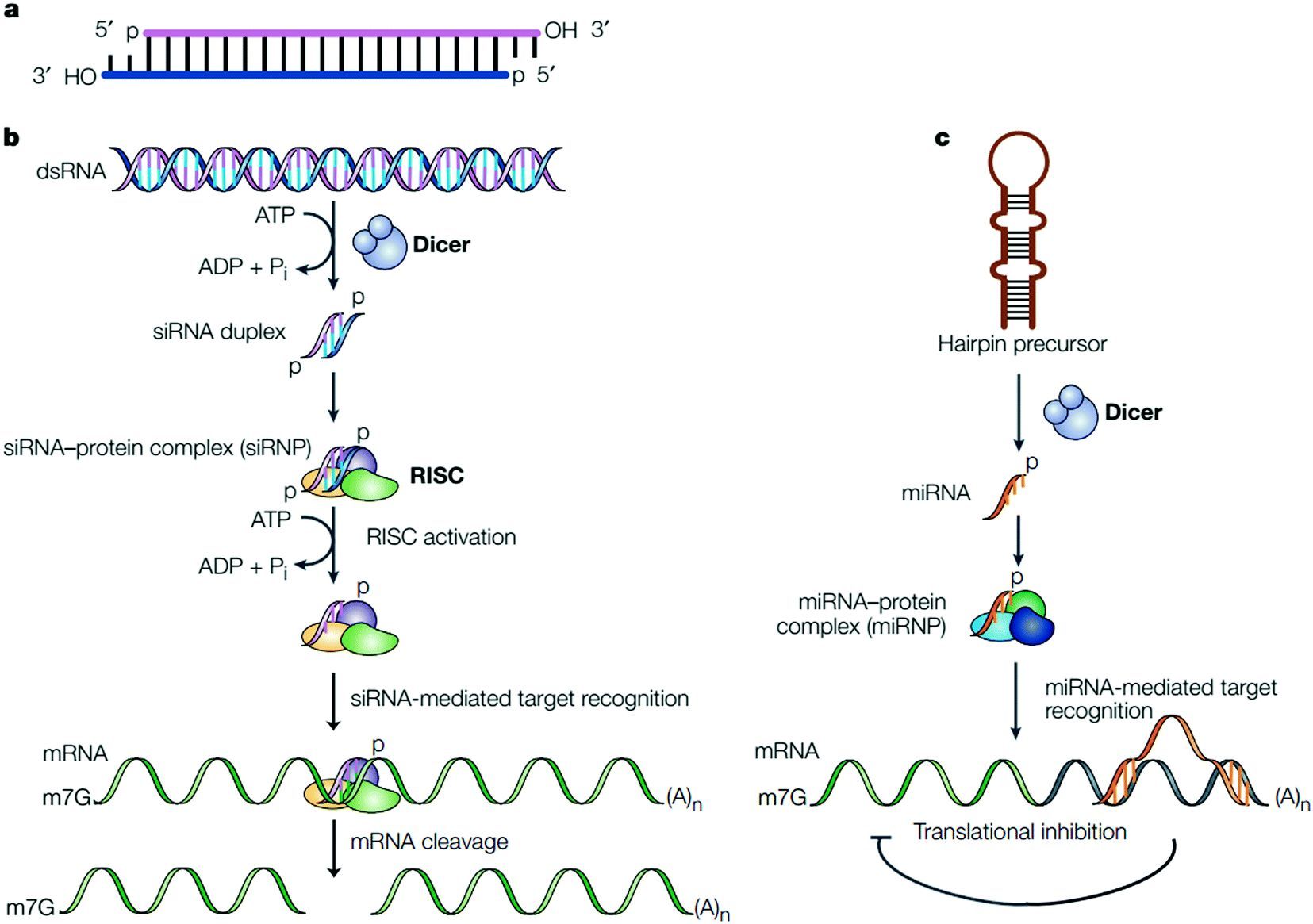
siRNA 不仅能引导 RISC 切割同源单链 mRNA,而且可作为引物与靶 RNA 结合并在 RNA 聚合酶(RNA-dependent RNA polymerase, RdRP)作用下合成更多新的 dsRNA,新合成的 dsRNA 再由 Dicer 切割产生大量次级 siRNA,经过若干次的合成-切割循环,RNAi 的作用不断放大,最终将靶 mRNA 完全降解。
siRNA 设计的原则
- 从 mRNA 的 AUG 起始密码开始,寻找”AA”二连序列,并记下其 3’ 端的 19 个碱基序列,作为潜在的 siRNA 靶位点。有研究结果显示 GC 含量在 30%-50% 左右的 siRNA 要比那些 GC 含量偏高的更为有效。Tuschl 等建议在设计 siRNA 时不要针对 5‘ 和 3’ 端的非编码区(untranslated regions, UTRs),原因是这些地方有丰富的调控蛋白结合区域,而这些 UTR 结合蛋白或者翻译起始复合物可能会影响 siRNA 核酸内切酶复合物结合 mRNA 从而影响 siRNA 的效果
- 将潜在的序列和相应的基因组数据库(人或者小鼠,大鼠等等)进行比较,排除那些和其他编码序列 /EST 同源的序列,例如使用 BLAST(www.ncbi.com.nlm.gov/BLAST/)。
- 选出合适的目标序列进行合成。通常一个基因需要设计多个靶序列的 siRNA,以找到最有效的siRNA 序列。
合成步骤
- 合成一条或者多条高质量靶向目的基因的 siRNA
- 将 siRNA 转染进细胞,最后 siRNA 与目的基因结合从而达到基因沉默的目的
- 设置荧光转染对照组检测当前转染条件下细胞的转染率

